Publicatie datum: 15-12-2004
Sturen van de sortering van aardappelen
In opdracht van het HPA heeft praktijkonderzoek Plant & Omgeving een literatuurstudie uitgevoerd naar de sturing van de knolsortering, met de nadruk op resorptie van aangelegde knollen. In dit artikel worden de resultaten van deze studie kort weergegeven.
Een voorspelling van het aantal knollen die aan een aardappelplant zullen komen is, ruim voor de aanleg van deze knollen, onmogelijk.
Algemeen
De maatsortering van aardappelen, voor welke bestemming dan ook, roept in de praktijk steeds weer vragen op. Een lastige doelstelling voor pootgoedtelers en sommige consumptieaardappeltelers is het realiseren van een zo groot mogelijk aantal knollen en daardoor een fijne sortering. Er zijn verschillende redenen en omstandigheden waardoor het uiteindelijke aantal knollen kan tegengevallen Zo kan droogte na het poten oorzaak zijn van een geringe knolaanleg. In 2002 viel het pootgoedtelers op dat zij na een aanvankelijk goede knolzetting later nogal wat geresorbeerde knolletjes vonden; knolletjes tot 25 mm die weer leeggezogen werden en voor een deel wegrotten.
Duidelijk is dat naarmate de knolopbrengst per hectare hoger is, het gemiddelde knolgewicht hoger zal zijn en de fractie in de grove maten ook. Bij gelijke opbrengst leiden meer knollen tot een lager gemiddeld knolgewicht en een fijnere sortering (mits de knolvorm niet verandert).
Het aantal knollen wordt door veel factoren bepaald, waarvan een deel goed te beïnvloeden is en een ander deel niet of nauwelijks, zie figuur 1.
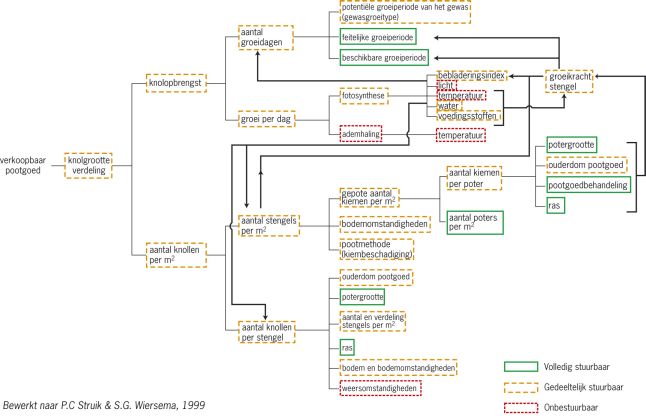
Knolvorming is dan ook een ingewikkeld proces. De eerste stap is de vorming van stolonen. Tijdens de groei van stolonen vindt onder gunstige omstandigheden knolinductie plaats; dat is het fysiologische signaal dat de stoloon aanzet tot knolvorming. Dit wordt gevolgd door knolinitiatie: het eerste opzwellen van de stolonen en het eerste zichtbare teken van knolaanleg, Daarna volgt de knolzetting: het zodanig uitgroeien van een knolaanlegsel dat het een gerede kans heeft om uit te groeien tot een oogstbare maat. In de fase erna vindt de knolgroei plaats tot de uiteindelijk te oogsten maat.
Gedurende de gehele knolgroeiperiode zijn alle knolvormingsprocessen in principe omkeerbaar. Dat wil zeggen dat als de omstandigheden voor knolinductie of knolgroei ongunstig zijn, knolvormingsprocessen kunnen stoppen, knolaanlegsels weer kunnen verdwijnen, en reeds gevormde knollen kunnen worden geresorbeerd. Onder bepaalde omstandigheden en bij bepaalde rassen is resorptie meer regel dan uitzondering. In die gevallen wordt het aantal oogstbare knollen niet alleen bepaald door de processen die inductie, initiatie en zetting beïnvloeden, maar ook door de processen die na de knolaanleg de knolgroei bepalen.
Het aantal knollen is afhankelijk van het aantal stengels en het aantal knollen per stengel. Deze beide worden in sterke mate beïnvloed door het ras.
Het aantal knollen per stengel is afhankelijk van het aantal gevormde stolonen per stengel en bij de stolonen kan weer onderscheid gemaakt worden naar de mate waarin deze vertakken.
In dit artikel willen we ons vooral concentreren op het aantal knolletjes dat gevormd wordt en het aantal knolletjes dat weer verdwijnt onder invloed van – vooral uitwendige – omstandigheden die zich tijdens de teelt voordoen en slechts gedeeltelijk stuurbaar zijn. Hierbij komen aan de orde; de stengelverdeling, het stikstofaanbod, de temperatuur en de hoeveelheid beschikbaar licht en water.
Stengelverdeling
Bij een gelijk aantal stengels per strekkende meter rug is bij een gelijkmatiger stengelverdeling over de rug het aantal knollen per stengel groter (Scholte, 1975). Bij grote poters staan de stengels meer in flinke pollen bijeen, terwijl bij kleine poters de stengels meer regelmatig over de rij zijn verdeeld. Dit is gunstig voor de lichtopvang van de grote bladeren onder aan de stengel en bevordert het aantal uitgroeiende knollen.
Een hoog stikstofaanbod remt de knolaanleg
In modelproeven in watercultures toonde Krauss (1985) aan dat de knolaanleg eenvoudig beïnvloed kon worden door de concentratie van stikstofionen te veranderen. Hij liet zien dat door een hoge concentratie stikstof de knolvorming volledig geremd kan worden of sterk verlaat, onder omstandigheden waarin bij lagere concentraties wel knolvorming optreedt. Als hij planten die met knolvorming begonnen waren, naar een oplossing verplaatste met een ‘excessief’ hoog N-niveau, dan stopte de knolvorming en begon de stoloongroei opnieuw. Zette hij deze vervolgens weer bij een laag stikstofniveau dan werd voor de tweede keer een knolletje gevormd (een soort doorwas, een ketting).
Vanuit veldproeven is het verlagende effect van hoge stikstofhoeveelheden op het aantal knollen ook bekend. Maar dit trad pas op bij hoeveelheden van 250 tot 300 kg zuivere stikstof per hectare bij het poten, in vergelijking met lagere stikstofhoeveelheden. Bij hoeveelheden van 250 kg en hoger was het aantal knollen bij de oogst lager dan bij wat lagere hoeveelheden stikstof. Onder de 250 kilo – en dus zeker bij stikstofniveaus die voor pootgoed gebruikelijk zijn – treedt dit effect niet of nauwelijks op.
Van de andere voedingselementen, zoals fosfaat en kali, is geen duidelijk verhogend effect bekend op het aantal knollen als ten minste de bodemtoestand minimaal voldoende is. De oude stelling dat een bemesting met fosfaat de knolzetting stimuleert, geldt dus alleen bij een (te) geringe fosfaattoestand van de bodem. En ook een fosfaatbladbemesting is onvoldoende bedrijfszeker als methode om het aantal knollen te verhogen (Rops & Floot, 1996).
Hoge temperaturen remmen de knolaanleg en knolgroei
Er zijn meerdere effecten van de temperatuur op de knolaanleg:
- Hoge temperaturen remmen de knolaanleg. Dit gebeurt zowel bij korte (10-12 uur) als bij lange dagen (15-18 uur). Het remmende effect is veel groter bij lange dagen zoals tijdens het groeiseizoen in Nederland dan bij korte dagen.
- Bij hogere temperaturen is de netto- fotosynthese lager, omdat er meer assimilaten door verademing verloren gaan. Dit komt doordat de ademhaling bij hogere temperatuur sneller toeneemt dan de fotosynthese.
- Bij hogere temperaturen (25-30 graden) gaat een groter deel van de assimilaten naar het loof en minder naar de knollen in vergelijking met lagere temperaturen (15-20 graden). Dit geldt ook voor de daglengte. Bij lange dagen gaat een groter deel naar het loof; bij korte dagen naar de knollen.
- Er is een scherp optimum voor de activiteit van zetmeelsynthetiserende enzymen. Dat wil zeggen dat bij hoge bodemtemperaturen van circa 30 graden het zetmeelgehalte van de knollen nauwelijks toeneemt, wat direct veroorzaakt wordt door de remming van de omzetting van suikers naar zetmeel (Krauss & Marschner, 1984).
Het is vooral de combinatie van weinig straling en een hoge temperatuur die het aantal knollen beperkt. Dit is voor een laboratoriumproef weergegeven in tabel 1 voor drie stralingshoeveelheden en drie combinaties van dag- en nachttemperaturen. Hierbij is het laagste stralingsniveau 3,4 MJ/m2/dag. Dit komt in Nederland in het veld nauwelijks voor. Op een donkere dag in Nederland is het in de periode juni-augustus al gauw 6 MJ/cm2/dag; op een heldere dag in augustus drie, vier keer zoveel (18-24 MJ/cm2/dag).
| Temperatuur °C | Straling, MJ / cm2 / dag | |||
| Dag | Nacht | 3,4 | 7,7 | 11,0 |
| 22 | 18 | 5,6 | 6,9 | 6,9 |
| 26 | 22 | 3,3 | 5,0 | 7,4 |
| 30 | 26 | 0,0 | 0,6 | 8,5 |
Uit deze tabel blijkt dat bij langdurig hoge temperaturen in combinatie met eveneens langdurig weinig licht minder knollen per plant worden aangelegd. Als we dit met Nederlandse omstandigheden in het veld vergelijken dan lijkt het niet waarschijnlijk dat er zodanige perioden optreden dat daardoor weinig knollen worden gevormd.
In een ander onderzoek is de invloed van de luchttemperatuur ten opzichte van de bodemtemperatuur nagegaan. Hierbij is de temperatuur van lucht en bodem dus apart geregeld. Het bleek dat hoge luchttemperaturen (hoog, 30-35 graden; laag, 17-27 graden) de knolinductie meer remden dan hoge bodemtemperaturen (Jackson, 1999).
Hoge bodemtemperaturen zorgen daarentegen, zoals eerder genoemd, dat de omzetting van suikers naar zetmeel wordt geremd en daarmee de toename van het drogestofgehalte. Daarnaast veroorzaken hoge bodemtemperaturen allerlei fysiologisch bepaalde vergroeiingen en misvormingen, zoals groeischeuren en doorwas.
Lage lichthoeveelheden remmen de knolaanleg
Het effect van lage lichthoeveelheden lijkt op dat van hoge temperaturen. Lagere temperaturen kunnen de effecten van minder straling op de knolaanleg compenseren zoals in tabel 1 te zien is. Het is dus vooral de combinatie van weinig straling en hoge temperatuur die het aantal knollen beperkt. Deze omstandigheden komen in de gematigde gebieden over langere perioden echter maar weinig voor.
Effect van daglengte op knolaanleg nauwelijks van praktisch belang
De knolaanleg vindt bij korte dagen van 12 uur eerder plaats dan bij langere dagen van 15 tot 18 uur. Dit betekent echter in de praktijk in Nederland weinig, waar de lichtperiode een vast gegeven is en in de periode mei/ juli varieert van 15 tot 18 uur.
Bij vroege droogte minder knollen
Haverkort et al. (1990) toonden aan dat bij vroege droogte, dat wil zeggen droogte tijdens de stoloonvorming, het aantal knollen per plant lager was ten opzichte van geen droogte en latere droogte. Dit is weergegeven in tabel 2. Latere droogte was droogte vanaf ongeveer een maand na opkomst, tijdens de knolaanleg, D2.
| Proef | Object | Behandeling | Aantal stengels per plant | aantal stolonen per stengel | knollen per stengel |
| 1 | M | geen droogte | 4.1 | 7.8 | 1.9 |
| D1 | droog van 7 tot 28 dagen na opkomst | 4.1 | 7.2 | 1.3 | |
| D2 | droog van 23 tot 48 dagen na opkomst | 4.1 | 7.3 | 2.0 | |
| 2 | M | geen droogte | 3.1 | 7.2 | 4.0 |
| D1 | droog van 10 tot 24 dagen na opkomst | 3.1 | 4.5 | 3.2 | |
| D2 | droog van 32 tot 43 dagen na opkomst | 3.1 | 6.8 | 4.7 |
De auteurs vergeleken dit met de waargenomen aantallen knollen in de “groeicurvebepalingen” bij het ras Bintje van de vroegere proefboerderij Feddemaheerd in Kloosterburen in de periode 1976 - 1988 en de hoeveelheid neerslag gedurende de eerste 40 dagen na poten. Ze vonden daarbij een duidelijke samenhang tussen hoeveelheid neerslag en aantal knollen per stengel: Meer neerslag in de eerste 40 dagen na poten ging samen met meer knollen per stengel (zie figuur 2).
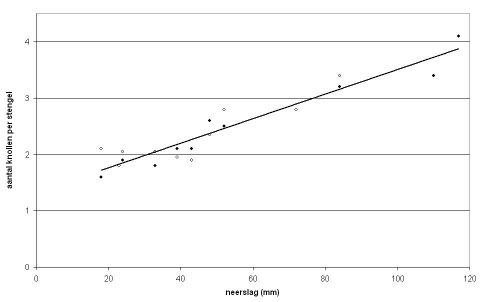
Op basis van deze resultaten verwachten de auteurs dat onder droge omstandigheden beregenen omstreeks de opkomst van het gewas het aantal knollen zal doen toenemen. Ook andere onderzoekers hebben de relatie tussen vocht en aantal knollen nagegaan en kwamen tot de conclusie dat heel vroege droogte, dus droogte voordat de knollen zijn aangelegd het meest beperkend was voor het knoltal. MacKerron & Jefferies (1986) concludeerden dat de aardappelplant voor wat betreft het aantal knollen maar een korte periode gevoelig is voor “waterstress”; twee, misschien drie weken. Droogte na knolaanleg had in hun kasproeven geen invloed op het aantal knollen. Volgens deze onderzoekers is het vooral belangrijk groeivertraging te voorkomen in de periode tussen opkomst en begin van de knolaanleg en daarmee beperking van het aantal uitgroeiende knolletjes.
Haverkort c.s. (1990) vonden bij de analyse van de “groeicurvebepalingen” geen verband tussen de hoeveelheid neerslag in de eerste 40 dagen na poten én het aantal stengels per plant. En hierbij maakte het geen verschil of voorgekiemd of niet voorgekiemd pootgoed werd gebruikt. Dit suggereert dat het aantal te vormen stengels vooral besloten ligt in de eigenschappen van het pootgoed en, afgezien van extremen, minder in de omstandigheden na het poten.
Sterke kluiterigheid van de ruggen geeft een slechtere aansluiting van de grond waardoor onder droge omstandigheden kiemen deels niet uitgroeien en zich dus minder stengels per plant ontwikkelen. Meer nog veroorzaken droge omstandigheden minder stolonen en knollen per stengel. Zo kon in een beregeningsproef in 1980, op proefboerderij ‘De Kandelaar’ bij Biddinghuizen (55% afslibbaar) op een grond met een heel kluiterige structuur het totale aantal knollen per m2 met behulp van beregenen worden verhoogd van 48 naar 74 en het aantal in de maat 28/45 mm van 28 naar 56 (Van Loon & Wassink, 1982). De eerste keer beregenen vond plaats toen pas enkele planten boven stonden. Door het beregenen nam in deze proef het aantal hoofdstengels per plant slechts toe van 3,5 naar 3,7. Kluiterige ruggen als gevolg van een slechte structuur versterken dus het effect van droge omstandigheden na het poten op het aantal knollen.
Effecten van fluctuaties
Tijdelijke, kortdurende veranderingen in de omstandigheden zijn in het wisselvalige Nederlandse klimaat gangbaar. Kijken we naar de variatie in weer van een bepaalde week in juni over de jaren heen dan kan die variatie enorm zijn. Vlak voor, tijdens en kort na de knolaanleg kan het aantal knollen sterk beïnvloed worden door toevallige fluctuaties in weer net tijdens die periode. Dit geldt vooral voor belangrijke factoren zoals temperatuur, lichtintensiteit, en vochtvoorziening. Daarom is in de praktijk de variatie tussen de jaren in knolzetting zo groot en daarom ook kunnen de verschillen binnen een jaar zo aanzienlijk zijn, bijvoorbeeld als gevolg van verschillen in pootdatum of regionale verschillen in neerslag.
Resorptie
Er is niet veel degelijk onderzoek gepubliceerd over resorptie van aangelegde knolletjes onder invloed van bijvoorbeeld perioden met erg lage lichtintensiteit of perioden met erg weinig regenval.
Volgens Ewing & Struik (1992) vond Burstall in Engeland in een proef een relatie tussen het aantal geresorbeerde knollen en beschaduwing. Bij continu 50% minder licht door beschaduwing werden meer aangelegde knolletjes geresorbeerd dan bij continu geen beschaduwing (zie figuur 3A). Toen in het jaar erop of alleen tot de knolaanleg of alleen tijdens de knolaanlegperiode of alleen na de knolaanlegperiode werd beschaduwd kon geen verschil met niet beschaduwen in hoeveelheid geresorbeerde knollen worden aangetoond (zie figuur 3B). Wel vond zij een lineair verband tussen het aantal aangelegde knollen en het aantal geresorbeerde knollen. Naarmate er meer knollen werden aangelegd, werden er ook meer geresorbeerd. Wel bleven er altijd meer knollen over naarmate er ook meer werden aangelegd.
(A) In 1984 waren er twee behandelingen: 50% beschaduwing gedurende het hele groeiseizoen en een controle zonder beschaduwing
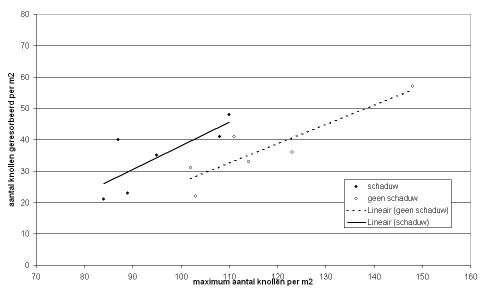
(B) In 1985 waren er 4 behandelingen: geen beschaduwing; 50% beschaduwing tot de knolaanleg; 50% beschaduwing tijdens de periode van knolaanleg; en 50% beschaduwing nadat de knolaanleg voorbij was, Ewing & Struik (1992)
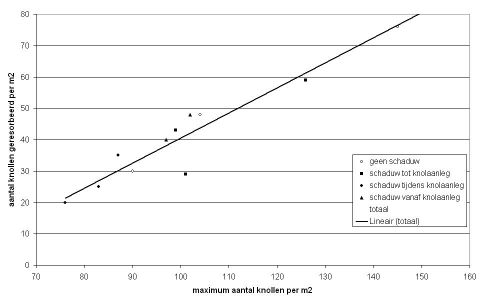
In 1979 en 1980 vonden Cho & Iritani (1983) in het noordwesten van de Verenigde Staten bij het ras Russet Burbank, dat naarmate later werd gepoot in de periode eind maart – half mei, er meer stengels en stolonen en knollen per plant werden gevormd. Ze zagen ook dat het aantal knollen vanaf juni juli sterk afnam, met 20 à 40%. Er was meer resorptie naarmate er later was gepoot. Uit de waargenomen cijfers blijkt ook dat waar de meeste knolletjes waren aangelegd uiteindelijk ook de meeste knollen overbleven. Ze noemden als oorzaak voor het hoge percentage resorptie, vooral in 1980, het grote aantal aangelegde knollen voor zo’n grofgroeiend ras als Russet Burbank, zie figuur 4. Er was een groot verschil in aantal knollen per plant tussen de beide jaren, maar duidelijk is wel dat waar meer knolletjes aangelegd zijn, er ook meer overblijven.
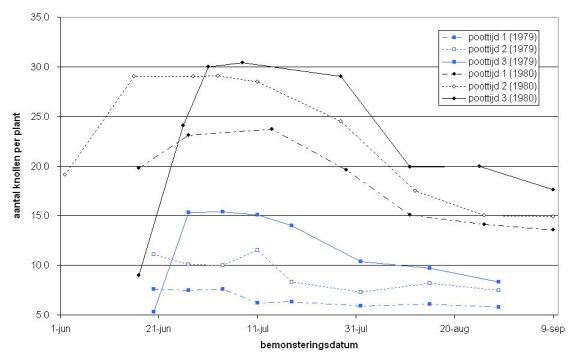
O’Brien et al. (1998) vonden in hun proeven dat aanhoudende erg droge omstandigheden in sommige proeven tot een verhoogd aantal geresorbeerde knollen leidden. Ewing & Struik (1992) stellen dat de beschikbaarheid van water één van de belangrijkste factoren lijkt te zijn die bepaalt welk deel van de knolaanlegsels uitgroeit tot grotere knollen.
In onderzoek in 1999 naar knolaanleg (Celis-Gamboa et al., 2003) met een groot aantal rassen op een zandgrond bij Wageningen, bleek dat van de grotere knolletjes (> 20 mm) 3 tot 12% weer verdween.
Ewing & Struik (1992) noemen als redenen waarom het proces van resorptie niet voldoende beschreven is, dat het ondergronds gebeurt en het probleem van het vaststellen wanneer je een knolaanlegsel een knolaanlegsel noemt en wanneer begin je van resorptie te spreken. Volgens Ewing & Struik (1992) is duidelijk dat er geen grens is waarna resorptie niet meer plaats kan vinden. Gegevens van Veerman (2001) bevestigen dit: gedurende het gehele groeiseizoen ontstaan er bij alle knolgewichten, naast de grote meerderheid van knollen waarbij het drogestofgehalte toeneemt ook steeds meer knollen met steeds lagere drogestofgehaltes. Zelfs snel groeiende knollen kunnen hun opgeslagen zetmeel weer kwijtraken door redistributie van suikers naar andere knollen. Zodra de balans tussen de aanvoer van suikers, de opslag van zetmeel en de afvoer van suikers negatief is, is er sprake van resorptie.
Discussie
Uit deze opsomming blijkt dat er onder normale omstandigheden maar heel beperkt mogelijkheden zijn om na het poten het aantal uitgroeiende knollen te beïnvloeden. Het belangrijkst is ervoor te zorgen dat de eerste weken na opkomst, tijdens de stoloonvorming, de grond voldoende vochtig is. Droge grond tijdens de eerste weken na opkomst beperkt het potentieel aantal aan te leggen knollen.
Voorts zijn er grote rasverschillen; het ene ras vormt veel meer knollen dan het andere. Maar dit verklaart zeker niet alles. Ook binnen een ras komen grote verschillen in aantal knollen voor.
De complexiteit van knolaanleg en knolgroei is buitengewoon groot. Dat maakt het zelfs achteraf vaak moeilijk precies aan te geven waarom de sortering geworden is, zoals die uiteindelijk is vastgesteld.
Beregenen tijdens de knolaanleg als maatregel ter beperking van gewone schurft is gangbaar, maar tijdens de stoloonvorming niet. Toch zijn er aanwijzingen dat hierdoor het aantal knollen kan worden verhoogd.
Door middel van onderzoek zou kunnen worden nagegaan in hoeverre het vochtig houden van de grond tussen opkomst en knolaanleg, en bij welke rassen, een effectieve maatregel is om de hoeveelheid aangelegde knolletjes te verhogen en daarmee ook meer knollen in de economisch interessante maten te oogsten.
Conclusie
Een voorspelling vooraf van het aantal knollen dat uitgroeit, is niet te maken. Wel hangt de maatsortering van aardappelen in sterke mate af van het aantal aangelegde stolonen en knolletjes. Voldoende vochtige omstandigheden tussen opkomst en knolaanleg zijn gunstig om veel stolonen en daarmee veel knollen per stengel te krijgen. De mate waarin dit bij de verschillende rassen werkt, is onvoldoende bekend. De invloed van de stikstofhoeveelheid, de temperatuur, de lichthoeveelheid, de daglengte en kluiterigheid op het uiteindelijke aantal knollen is onder normale omstandigheden heel beperkt.
Uit de beperkte kennis over resorptie blijkt dat er meer resorptie plaatsvindt naarmate er meer knollen zijn aangelegd. Toch zal het uiteindelijke aantal knollen bijna altijd hoger zijn naarmate meer knollen zijn aangelegd. Zowel het ras als de omstandigheden (vocht- en lichtgebrek) lijken van invloed op de mate waarin resorptie optreedt.
Literatuur
Celis-Gamboa, C. , Struik, P. C., Jacobsen, E., Visser, R. G. F., 2003. Temporal dynamics of tuber formation and related processes in a crossing population of potato (Solanum tuberosum). Ann. Appl. Biol. 143: 175-186.
Cho, J. L., Iritani, W. M., 1983. Comparison of growth and yield parameters of Russet Burbank for a two-year period. Am. Pot. J. 60: 569-576.
Ewing, E. E., Struik, P. C., 1992. Tuber formation in potato: induction, initiation, and growth. Hort. Reviews 14: 89-198.
Jackson S. D., 1999. Multiple signaling pathways control tuber induction in potato. Plant Physiology 119: 1-8.
Haverkort, A. J., Waart, M. v. d., Bodlaender, K. B. A., 1990. The effect of early drought stress on numbers of tubers and stolons of potato in controlled and field conditions. Pot. Research 33: 89-96.
Krauss, A., 1985. Interaction of nitrogen nutrition, phytohormones, and tuberisation. In Potato Physiology (Ed. P. H. Li). Pp. 209-230. Orlando FL: Academic Press.
Krauss, A & H. Marschner, 1984. Growth rate and carbohydrate metabolism of potato tubers exposed to high temperatures, Potato Research 27; 297-303.
Loon, C. D. v., Wassink, E. G., 1982. Bij de pootgoedteelt streven naar veel knollen. Bedrijfsont. 13: 579-581.
MacKerron, D. K. L., Jefferies, R. A., 1986. The influence of early soil moisture stress on tuber numbers in potato. Pot. Research 29: 299-312.
Menzel, C. M., 1985. Tuberisation in potato at high temperatures: interaction between temperature and irradiance. Annals of Botany 55: 565-568.
O’Brien, P. J., Allen, E. J., Firman, D. M., 1998. A review of some studies into tuber initiation in potato (Solanum tuberosum) crops. J. Agr. Sci, 130: 251-270.
Rops, A & H.W.G. Floot, 1996. Invloed van fosfaat-bladbemesting op de opbrengst en het knoltal van pootaardappelen. Jaarboek 1995/1996 PAGV, publikatie nr. 81A: p.41-44.
Scholte, K., 1975. De invloed van de stengeldichtheid op de produktie en sortering van consumptie-aardappelen. Bedrijfsontw. 6, 9: p. 731-735.
Struik, P. C., Wiersema, S. G., 1999. Seed potato technology. Wageningen Pers, Wageningen, pp 383.
Veerman, A., 2001. Variatie in knolkwaliteit tussen en binnen partijen van consumptieaardappelrassen. PhD Thesis Wageningen University, pp. 253.
Waart, M. v. d., 1991. Een vroege beregening verhoogt het aantal knollen. Aardappelwereld, 4: 31-33.