Publicatie datum: 01-07-2000
Pallida-resistente rassen onvermoeibaar?
Met de huidige teelt van E-resistente zetmeelaardappelrassen is het aardappelmoeheidsprobleem in Noordoost-Nederland voorlopig naar de achtergrond verdwenen. Om te toetsen hoe lang dit zorgeloze bestaan voort zal duren, heeft het HLB vanaf 1991 meerjarige veldproeven op zes locaties uitgevoerd, waarbij het effect van de teelt van de rassen Elles en Seresta op de pathogeniteit (= vermeerdering op resistent ras) van G. pallida-populaties is onderzocht. Daarbij is ook het effect van onderlinge afwisseling en afwisseling met Elkana meegenomen. Een kasproef met dezelfde G. pallida-populaties is in 1993 opgezet om de veldresultaten te onderbouwen.
In beide proeven is een verhoogde vermeerdering van de aaltjespopulaties als gevolg van de teelt van D- en E-resistente rassen waargenomen. Met ander woorden: er was sprake van uitselectie. De praktijk moet bij de teelt van E-resistente rassen in de (verre) toekomst rekening houden met stijgende besmettingsniveaus als gevolg van uitselectie naar pathogenere populaties.Voorlopig zullen bij de teelt van E-resistente rassen de besmettingsniveaus onder de schadedrempel blijven. Het afwisselen van E-resistente rassen met vatbare of D-resistente rassen kan selectie vertragen. Het middel is echter erger dan de kwaal, omdat door afwisseling de besmettingsniveaus voor een volgende teelt bij voorbaat boven de schadedrempel zullen uitstijgen.
Inleiding
Op veel bedrijven wordt de laatste tijd het aardappelmoeheidsprobleem met succes bestreden door de teelt van zetmeelaardappelrassen met hoge resistentie tegen het aardappelcystenaaltje Globodera pallida. Uit praktijkgegevens blijkt dat het AM-probleem bij deze teelt voorlopig van de baan is. Om er achter te komen hoe lang deze situatie standhoudt en wat de invloed van rassenkeuze hierop kan zijn, is het HLB mede op initiatief van het PAV begin jaren '90 met een grootschalig veld- en kasexperiment begonnen. De opzet was vrij uniek, omdat de proef zowel in het veld als in de kas (met de cysten uit het veld) is uitgevoerd. Bovendien is voor het eerst een hoogresistent ras als Seresta bij het onderzoek betrokken. De resultaten van het onderzoek geven duidelijkheid over het effect van de teelt van dergelijke rassen op de langere termijn.
Veld- en kasproef
In de periode 1991-1992 zijn door het HLB zes grootschalige proefvelden aangelegd, waar tot het jaar 2002 ieder jaar aardappelen zijn geteeld (tabel 1). Op de proefvelden is het besmettingsverloop van het aardappelcysteaaltje onder monocultuur nauwkeurig gevolgd. Er zijn verschillende locaties gekozen, in de verwachting dat daardoor ook verschillende G. pallida-populaties werden getoetst.
| Perceelscode | Locatie | Grondsoort | Organische stofgehalte (%) | pH | Eerste proefjaar |
| USK | Rolde (Proefboerderij Kooijenburg) | zand | 3,0 | 5,0 | 1992 |
| USS | Annerveenschekanaal | dal | 33,4 | 4,6 | 1992 |
| USV | Valthermond (proefboerderij 't Kompas) | dal | 16,9 | 4,7 | 1992 |
| USE | Vroomshoop | zand | 7,5 | 4,4 | 1993 |
| USH | Buinen | zand | 3,7 | 4,9 | 1993 |
| USP | Rheederveld | dal | 11,2 | 5,1 | 1993 |
Een overzicht van de uitgevoerde rasafwiiseling is weergegeven in tabel 2.
| Object | Teelt | Resistentie |
| A | Seresta - Seresta | Ro1,2,3,4 Pa2,3 |
| B | Elles - Elles | Ro1,2,3,4 Pa2 |
| C | Elkana - Elkana | Ro1,2,3,4 |
| D | Elkana - Seresta | |
| E | Elles - Seresta | |
| F | Elles - Elkana |
De rassen Elkana, Elles en Seresta zijn continu en afwisselend geteeld. De teelt van Elles en Seresta was bedoeld om selectiedruk op de aaltjespopulatie uit te oefenen en de gevolgen daarvan te onderzoeken. De rasafwisseling was bedoeld om mogelijke effecten daarvan op het uitselectieproces aan te kunnen tonen. Door de jaarlijkse teelt van Elles en Seresta loopt het onderzoek vóór op de praktijk (met name bij Seresta).
Parallel aan het veldonderzoek heeft het HLB samen met het toenmalige CPRO-DLO een kasproef uitgevoerd. De proef was bedoeld als onderbouwing van de resultaten uit het veld. Bovendien was het mogelijk om twee aardappelteelten per jaar uit te voeren, zodat het onderzoek meer vaart zou krijgen. De zes veldpopulaties zijn in de kas in potten vermeerderd op de rassen Irene, Elles en Seresta. Hiermee zijn per veldpopulatie drie G. pallida-lijnen gekweekt, waarvan één (vermeerdering op Irene) als onbeselecteerde en twee (vermeerdering op Elles en Seresta) als beselecteerde lijnen kunnen worden beschouwd. Na elke vermeerdering is onderzocht of de pathogeniteit van de beselecteerde lijnen was veranderd ten opzichte van de onbeselecteerde lijn.

Resultaten
Veldproef
In figuur 1 is het besmettingsverloop op de proefvelden bij de teelt van Elles en Seresta weergeven.
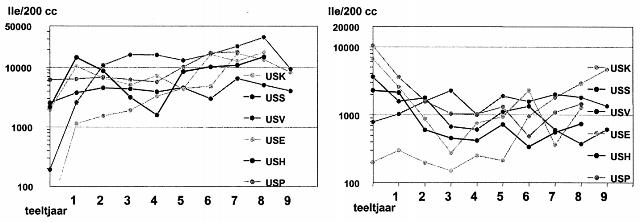
Uit de linker figuur blijkt dat bij de teelt van Elles bij een 1:1-teelt vaak schadelijke populatiedichtheden werden gemeten. Er is tussen een aantal locaties sprake van een duidelijk niveauverschil. Dit is enerzijds veroorzaakt door verschil in de samenstelling van de aaltjespopulaties, anderzijds door verschil in effectiviteit van het ontsmetten. Op het proefveld in Rolde (onderste lijn) bleef de besmetting zonder ingrijpen lange tijd vrij laag, terwijl op de andere proefvelden vrijwel ieder jaar moest worden ontsmet. Op het proefveld in Valthermond (bovenste lijn) zijn na een aantal teelten zeer hoge aaltjesdichtheden gemeten, mede door de geringe effectiviteit van het ontsmetten. De hoge dichtheden in de figuur maken in ieder geval duidelijk dat op de meeste locaties al een forse Pa3-populatie aanwezig was, die zich op Elles goed kon vermeerderen.
Uit de rechter figuur blijkt dat de teelt van Seresta over het algemeen een sterke daling van de besmettingsniveaus tot gevolg had. Ook bij deze teelt toonden de populaties onderlinge verschillen. Op het proefveld in Valthermond bijvoorbeeld bleef het besmettingsniveau erg hoog. Wat de meeste populaties wel gemeen hebben is de kentering in de dalende lijn na zo'n vier Seresta-teelten. Opvallend daarbij is de stijgende lijn van de populatie die in het begin erg mild was.
De vermeerdering op Elkana (van de uitgangspopulatie, die steeds op dit ras is vermeerderd) is in de loop van de jaren nauwelijks veranderd. De vermeerderingslijnen van de eerste vier en de laatste twee teelten liggen bij dit ras dicht bij elkaar. Van nadelige jaarsinvloeden is dus weinig sprake geweest. Hieruit kan worden opgemaakt dat de vermeerdering op Elles en Seresta óók niet nadelig is verstoord. Eventuele veranderingen bij de teelt van Elles en Seresta kunnen worden toegeschreven aan verandering van de pathogeniteit van de populaties.
Uit figuur 1 blijkt dat bij de continuteelt van Elles gemiddeld een geringe verhoging is opgetreden in pathogeniteit. Bij Seresta is de pathogeniteit gemiddeld sterk gestegen. Bij Seresta heeft deze stijging echter nog geen schadelijke besmettingsniveaus tot gevolg gehad (met uitzondering van USK).
Uit figuur 2 kan worden opgemaakt of de herhaalde teelt van Elles of Seresta ook een verandering in pathogeniteit heeft teweeg gebracht.
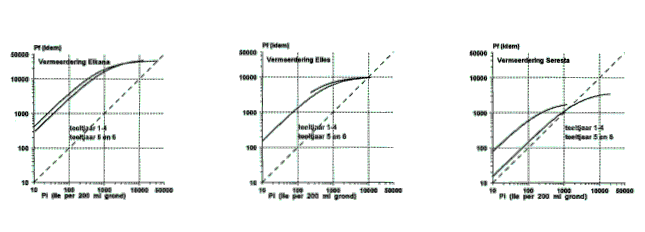
Uit de eerste waarnemingen bij rasafwisseling ontstaat de indruk dat afwisseling van rassen binnen het bouwplan in sommige gevallen effect heeft op de pathogeniteit van de populaties, maar de resultaten wijzen niet altijd in dezelfde richting. Afwisseling met Elkana had bijvoorbeeld even vaak een verlagende als een verhogende werking op de pathogeniteit voor Elles. Afwisseling met Seresta had bij twee van de getoetste populaties geen effect en bij twee populaties een verlagend effect op de pathogeniteit voor Elles. Rasafwisseling met Elkana of Elles had bij een aantal populaties ook een verlagende werking op de pathogeniteit voor Seresta. Bij de meest pathogene populatie (Valthermond) had deze rasafwisseling eerder een verhogende werking op de pathogeniteit voor Seresta. Los van het feit dat er nu nog geen uitsluitsel kan worden gegeven over de effecten van rasafwisseling op de pathogeniteit, is wel zeker dat in bouwplanverband afwisseling tussen hoogresistent en vatbaar leidt tot hogere besmettingsniveaus en meer kans op schade. Afwisseling tussen hoogresistente rassen (van verschillende herkomst) zou daar mogelijk beter op aansluiten.
Kasproef
De resultaten van de kasproef waren veel extremer dan van de veldproef. Bij alle populaties in de kas was na vier Elles-teelten de pathogeniteit van de beselecteerde lijn hoger ten opzichte van de onbeselecteerde lijn. De populatie uit Rolde (onderste lijn in figuur 1) gaf ook een duidelijke stijging te zien. De vermeerdering op Elles steeg bij deze populatie na vier Elles-teelten van vier naar tien (bij een beginbesmetting van vier levende larven en eieren per gram grond). Het effect van vier Elles-teelten werkte gemiddeld even sterk door op de pathogeniteit voor Elles als op die voor Seresta (de pathogeniteit voor beide rassen lag gemiddeld drie keer zo hoog ten opzichte van de onbeselecteerde populatie). Bij alle getoetste populaties is na drie vermeerderingen op Seresta de pathogeniteit voor Seresta sterk gestegen. Bij een populatie uit Rheederveld was na drie Seresta-teelten de vermeerdering op Seresta gestegen van 0,3 naar 3. Bij een populatie uit Buinen was de vermeerdering na drie teelten gestegen van 0,1 naar 1,9.

Conclusie
Bij herhaaldelijk telen van E-resistente rassen zal de praktijk in de (verre) toekomst weer te maken kunnen krijgen met stijgende besmettingsniveaus. Uit de resultaten van de veld- en de kasproef blijkt dat bij aaltjespopulaties in Nederland de aanleg aanwezig is om een E-resistent ras als Seresta vatbaarder te maken. Uit de resultaten in het veld blijkt dat de verschuiving op de meeste velden traag verloopt, maar dat het ook mis kan gaan.
De snelheid van het uitselectieproces is afhankelijk van de teeltfrequentie van het E-resistente ras en de samenstelling van de aaltjespopulatie op het perceel. Afwisseling met vatbare rassen kan het proces mogelijk vertragen, maar leidt wel meteen tot onaanvaardvare besmettingsniveaus, zodat het uiteindelijke doel van deze afwisseling (het laag houden van de besmetting) niet wordt gehaald. Afwisseling van E-resistente rassen past wat dit betreft beter in deze strategie. In de meeste gevallen is permanent overschakelen op E-resistente rassen, waarbij met regelmaat de eindbesmetting in de gaten wordt gehouden, de beste remedie.
Begin 2003 wordt een kasproef uitgevoerd, waarbij alle veldpopulaties worden getoetst op pathogeniteit, zodat met name over het effect van rasafwisseling meer duidelijkheid gegeven kan worden.Voortzetting van een tweetal veldproeven zal aan moeten tonen tot welke niveaus de populaties zullen stijgen bij continue inzet van E-resistente rassen.